Original Article
The Role of Pharyngeal Collapse in Nocturnal Homeostasis:
Exploring the Benefits and Costs of a Vastly Prevalent Phenomenon under an Evolutionary Framework
Jared Reser 06/05/04
Please send all inquiries to:
jared@jaredreser.com
Abstract
The present article will explore how the collapse of the pharyngeal airway during sleep, a phenomenon currently
identified as an indicator of disease, may confer specific homeostatic benefits. More specifically, we will consider
how pathologically large size of the fleshy structures in the pharyngeal airway, along with relative hypotonicity of
the pharyngeal musculature may have provided our ancestors a selective advantage by enabling them to reduce
nocturnal, respiratory heat loss. When the pharynx is collapsed, the diameter of the pharyngeal airway
decreases, allowing increased intermolecular collisions between inspired air and the epithelial walls of the upper
airway. An increase in the number of collisions should facilitate the transference of both warmth and humidity to
inspired air before it reaches the more sensitive lower airway resulting in the maintenance of internal, core
temperature and alveolar heat retention. By inducing increased muscular activity in the diaphragm and by
mechanically stimulating specific pharyngeal structures, pharyngeal collapse might provide other homeostatic
benefits as well. In light of the debilitating disorders associated with pharyngeal collapse, the absence of a
disease state secondary to surgical correction, the high proclivity for collapse documented in pregnant mothers,
and the functional similarities between collapse and reflex bronchoconstriction, it appears that the phenomenon is
explicable under evolutionary medicinal theory.
Keywords
bronchoconstriction, genetic epidemiology, hypotonia, mean free path, obstructive sleep apnea syndrome,
pharyngeal collapse, pulmonary mechanics
Introduction
The collapse of the pharyngeal airway, a physiological occurrence that accompanies sleep, is known to affect all
people. The degree of collapse experienced by an individual is determined by his or her genetic vulnerability and
can be placed along a spectrum from healthy breathing to apnea and disease. In susceptible individuals,
pharyngeal collapse presents as obstructive sleep apnea syndrome (OSAS), and upper airway resistance
syndrome (UARS), two widely prevalent, relatively pernicious disorders.
Using a paradigm constructed by Williams and Nesse (1991) this article will attempt to analyze pharyngeal
collapse using evolutionary medicinal theory. In order to show that a trait may be a form of evolutionary medicine,
under this framework, it is necessary to provide evidence that the trait is relatively prevalent, that it is heritable
and that susceptibility varies within populations. It is also necessary to show how the genes responsible for the
trait might have been naturally selected. For example, many heritable traits involve a tradeoff or compromise,
and, when analyzing these, it is necessary to provide evidence showing that the decrement in reproductive
success due to associated health risks can be mitigated by associated health benefits (Williams & Nesse 1991).
Williams and Nesse have identified many such “pathological” conditions such as anxiety, cough, diarrhea, fever,
inflammation, pain, sneezing and vomiting and have helped to show that they are actually evolved defenses
(1998).
This article will examine how pharyngeal collapse might create a net fitness advantage by playing a passive but
significant role in the maintenance of nocturnal homeostasis. Today, the costs of collapse are well documented
but the defensive manifestations may be hidden because of compensatory aspects of our modern environment
(ie. insulation and central heating). Providing an evolutionary explanation for susceptibility to collapse may help
explain the high prevalence in human populations and may inform both medical and gene therapy. Before we
consider the ultimate or evolutionary cause of pharyngeal collapse we must first look at the proximate,
pathophysiological causes. Because the present hypothesis is in part predicated on pathophysiological data, the
provision of background information describing the etiology, symptoms, and methods of intervention is in order.
Etiology of Pharyngeal Collapse
The discussion of etiology motivates us to focus on two causal factors, namely the collapse caused by hypotonia
(relaxation) in pharyngeal musculature, and the obstruction caused by specific structures within the pharynx.
Pharyngeal collapse arises when the airway that allows easy and normal breathing becomes blocked. When a
person is awake pharyngeal dilator muscles maintain a clearly defined, open pathway to the lungs. These
muscles disallow specific, membranous structures within the airway from interfering with inhalation. When a
person is asleep, however, these muscles become hypotonic; they relax and may tend to collapse (Horner 1996).
It is well accepted that the nocturnal collapse of pharyngeal musculature is responsible for both the increase in the
degree of obstruction caused by specific pharyngeal structures, and the restriction of the flow of air through the
upper and pharyngeal airways (Horner et al. 1994; Wheatley et al. 1993).
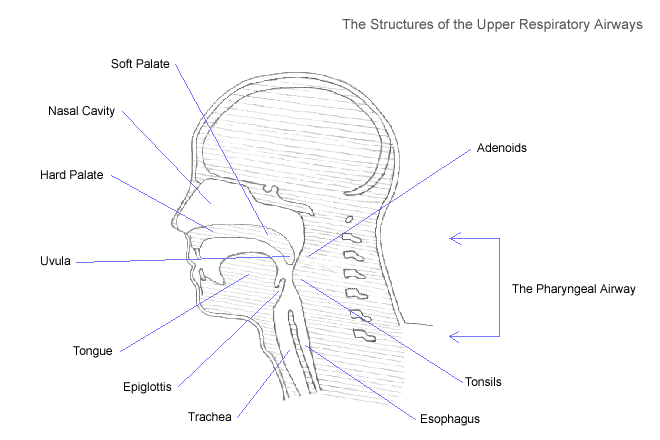
The sound associated with severe pharyngeal obstruction, snoring, occurs when certain structures within the
pharyngeal airway vibrate excessively as air is pulled through the narrow opening, toward the lungs. Any of the
membranous structures that lack cartilaginous support, including the uvula, adenoids, soft palate, pharyngeal
walls, and tonsillar pillars, are predisposed to vibration (Crumley et al. 1987). The soft palate (the upper surface
of the mouth that separates the oral and nasal cavities) and the uvula (the small piece of soft tissue that hangs
from the soft palate) are relatively large and fleshy, and can obstruct the upper airway completely. When these
structures obstruct the upper airway for even a short period of time they may cause discomfort or morbidity.
Sequelae and Associated Disorders
The complications associated with pharyngeal collapse are numerous and they vary in severity. A review of these
complications allows insight into how such a potent dysfunction might have seriously jeopardized the evolutionary
fitness of our ancestors.
Repetitive and severe obstruction of the pharyngeal airway results in the upper airway resistance syndrome
(UARS), a form of sleep disordered breathing in which resistance to airflow leads to brief arousals from sleep
thereby causing daytime somnolence (weariness) (Zielinski et al. 1999). Many UARS sufferers experience
episodes of severe obstruction lasting from a few seconds to a full minute. These episodes take two forms:
hypopnea (periods of reduced breathing) and apnea (cessation of breathing). The related disorder, obstructive
sleep apnea syndrome (OSAS), often results in a loss of oxygen which can create serious disturbances in health.
The cessation of breath caused by obstruction can induce hypoxemia, a state defined by insufficient oxygen in the
blood (oxyhemoglobin desaturation). It has been estimated that 24% of men between the ages of 30 and 60
suffer from apnea induced hypoxemia (Young et al. 1997). Painful cluster headaches precipitated by hypoxemia
are also strongly associated with OSAS (Kudrow and Kudrow 1990, 1993).
Prolonged hypoxemia often results in hypoxia, a deficiency in the amount of oxygen reaching body tissues.
Hypoxia induced by OSAS is known to cause tissue damage in various regions of the body (Jennum et al. 1993),
including the brain. It has been suggested that hypoxia caused by OSAS is linked with hypertension (Young et al.
1997), heart disease (Lugaresi et al. 1980), cerebrovascular disease (Neau et al. 1995), and various other
cardiovascular diseases (Lavie et al. 2001). Recognition of the dangers associated with OSAS within the
literature is a cause of increasing public interest and concern (Shin et al. 2003).
Medical Intervention
An analysis of the methods of medical intervention reveals that the causal pharyngeal structures can be reduced
surgically with virtually no negative consequence. There are several methods of intervention aimed at mitigating
pharyngeal obstruction and they range from oral devices to surgery. The most successful way to stop habitual
snoring is the surgical removal of portions of the soft palate (Krespi et al. 1994). Doctors that practice head and
neck surgery, otolaryngology, perform such operations not as a last resort, but as a relatively successful method
of medical intervention (Utley et al. 1997). Such surgical procedures include uvulopalatopharyngoplasty (UPPP),
Laser-Assisted Uvuloplasty (LAUP) and palatal reduction by radio frequency ablation (somnoplasty). Larger
populations of patients must be analyzed before surgery can be recommended as the primary form of therapy;
however, there are currently no persisting, negative complications associated with the surgical reduction of the
soft palate (Powell et al. 1998). Because the reduction of the soft palate poses no threat to modern day patients,
an evolutionary explanation for the wide prevalence of its excessive size should prove informative.
The Prevalence of Pharyngeal Collapse
Pharyngeal collapse is a phenomenon that affects all people and it can result in various disorders that are highly
prevalent in human populations around the world. Studies have demonstrated that the prevalence of OSAS could
vary between 3.2% and 12.1% in children (Corbo et al. 1989; Anuntaseree et al. 2001), and between 5% and 40%
in adults (Bearpark et al. 1995; Cirignotta et al. 1989; Fitzpatrick et al. 1993; Redline et al. 1997). Also elderly
populations are at significantly higher risk than younger populations (Young et al. 1993). Snoring is also very
common in the general population; 35–45% of men and 15–28% of women report that they snore habitually
(Ohayon et al. 1997; Young et al. 1993). The elderly, adults, children and even infants (Brouillette et al. 1982)
experience pharyngeal collapse and the associated sequelae. The presence of collapse throughout the
maturational timeline is indicative of its value as a protective mechanism.
Like most inherited traits, and protective adaptations, a person’s proclivity for pharyngeal collapse is genetic
(Davies et al. 1996; Silverberg et al. 1997). Many researchers consider it likely that inherited, familial factors such
as upper airway structure and neural control of the upper airway muscles interact to produce relative proclivity for
OSAS (Redline et al. 2002). Because environmental factors can vary greatly from area to area, the relative
distribution of most protective traits is known to vary widely, even between members of the same species. As
might be expected, a large amount of variance exists with regard to the severity of pharyngeal collapse within the
human species. Wide ethnic variations (Redline et al. 1997) in the prevalence of habitual snoring might result from
differences in definitions and methodology, but more importantly it may stem from differences in population
genetics. In fact both the incidence of habitual snoring and the number of studies describing OSAS appear to be
far less prevalent in Asia (Gau et al. 1995; Hui et al. 1999).
Hypothesis: Pharyngeal Collapse May Reduce Respiratory Heat Loss
If the risks associated with pharyngeal collapse make it a potentially enervating and morbid activity, how then
could it have become so ubiquitous? Why would the human body so dramatically decrease autonomic, muscular
support to the throat during rest, and why would such large, fleshy structures exist in an area so vital to respiration
and health? Despite the scarcity of related research the present article will attempt to characterize pharyngeal
collapse as a mechanism, much like other respiratory mechanisms, that promotes the maintenance of
homeostasis by regulating respiratory heat retention. Collapse occurs exclusively during sleep- this may be a
functional response to the cold exposure that accompanies sleep related processes.
Factors that Expose Sleeping Animals to the Cold
There are at least two factors that expose diurnal, sleeping animals to the cold. When an animal is sleeping its
skeletal muscles are quiescent, especially during deep sleep. A lack of muscular exertion is a common cause of
reduction in internal temperature. Also, it is widely recognized that the temperature of nighttime air is relatively low
compared to daytime air. This temperature difference exposes diurnal animals to the cold when they are the most
vulnerable. For these two reasons diurnal animals are particularly susceptible to heat loss during sleep.
The sleeping mammal’s respiratory tract is a prime candidate for the cooling process because it contains many
square inches of wet, soft tissue that is exposed to rapidly moving, dry air for many hours at a time (Eagan 1963).
As each new breath of air is inhaled, water molecules evaporate from the throat and lungs as part of an
endothermic process, and a potentially large amount of body heat can be lost (Ferrus et al. 1980). For example,
dogs utilize the respiratory tract as their primary means of dissipating excessive heat (Iscoe et al. 1983).
Humans are susceptible to substantial changes in external temperatures and will die if their core, internal
temperature drifts more than a few degrees above or below 37° C (Lloyd 1994). It is fortunate that humans, as
homeothermic mammals, are thermoregulators. They utilize specific homeostatic mechanisms to minimize the heat
lost to the external environment. Such mechanisms include vasoconstriction, concurrent heat exchange,
metabolic regulation and shivering. These methods of heat conservation focus on maintaining the core, internal
temperature at the expense of heat loss within the extremities. Heat loss arising from respiration therefore can be
very potent because it conducts heat away from the lungs and thorax, the very center of the body. If it is true that
nocturnal, sleeping processes are more susceptible to heat loss than diurnal, wakeful processes, and it is true
that pharyngeal collapse is a mechanism involved in minimizing the rate of heat exchange between the respiratory
system and the surrounding environment, then the involvement of collapse as an exclusively nocturnal mechanism
is quite appropriate.
The Importance of Heating Inspired Air
The respiratory airways both heat and humidify inspired air before it reaches the alveoli, the final destination in
the respiratory tract (Millar et al. 1965). Researchers do not seem to have reached a consensus regarding the
benefits of maintaining the temperature of the lower airways and alveoli, but they do seem to agree on its
importance. McFadden reported that the inhalation of dry, cold air increases cardiovascular stress and
decreases pulmonary function (1983). Furthermore, certain reflexive measures taken by the respiratory system
when it is exposed to cold air have led researchers to believe that cold air is deleterious to respiratory health and
functionality (McFadden 1983). The exposure of the airways to cold air is consistently accompanied by reflex
bronchoconstriction, vasoconstriction and the release of histamines (Doyle et al. 1994; Geller et al. 1988). These
responses are regarded by researchers to be protective (O’Cain et al. 1980).
Contact of the skin with cold air is well known to stimulate thermally sensitive receptors that produce reflexive
bronchoconstriction (Doyle et al. 1994). The constriction of the passageways within the tracheo-bronchial tree
(the branches leading to the alveoli) in response to cold air is a process that diminishes susceptibility to heat and
humidity loss (O’Cain et al. 1980). In other words, the heat and humidity retention that I am ascribing to the
collapse of the pharyngeal airway has been previously ascribed to the collapse of the tracheo-bronchial tree; the
two mechanisms are physically and functionally analogous. The benefits conferred by reflex bronchoconstricion
can arise when the animal is both asleep and awake. Unlike reflex bronchoconstriction, pharyngeal collapse
responds specifically to the heat loss associated with nocturnal processes. A careful, physical analysis shows that
collapse should increase respiratory heat conservation by increasing the rate of molecular collisions within the
pharyngeal airway.
The Collapse of the Pharynx Should Result in the Warming of Inspired Air
Pharyngeal collapse should help to warm inhaled air before it reaches the lungs by creating a “narrower tube” for
air to travel through. This narrow tube should also provide other benefits including the retention of water vapor,
and the filtration of inspired pathogens. It should seem conceivable that a broader airway should not prove to be
as effective as a narrower airway in achieving these benefits. For clarification, let us turn to an academically
popular, thermodynamic system that is structurally analogous to the collapsible, pharyngeal airway.
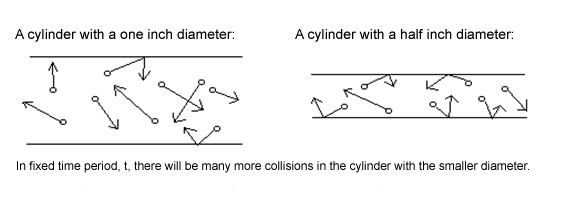
From a physical perspective a “narrowing airway” is a cylinder that exhibits a variance in the distance between its
walls. The number of times that a gas molecule within such a cylinder will collide with the walls of its container is a
function of the distance between the walls (Wark et al. 1999). As the walls draw nearer, the average distance that
the molecules will travel before they collide with one of the walls decreases. In other words, a reduction in the
distance between walls results in an increase in the number of collisions per time period (Serway 1990).
The number of collisions is very important because it determines how much kinetic energy will be transferred from
the walls to the air molecules by conduction (Serway 1990). Knowledge of the corollaries of this physical model
allows us to conclude that a narrower airway will create more collisions between the cold air molecules and the
warm epithelial tissues of the pharyngeal airway thereby increasing the temperature of inspired air before it
reaches the alveoli.
The Conservation of Water Vapor
Considerable water loss takes place through exhalation (Ferrus et al. 1980), and by narrowing the airway, more
water vapor should be conserved. As mentioned before, a decrease in the distance between pharyngeal walls
produces more molecule-wall collisions. The kinetic theory of gasses predicts that as the number of collisions
increases, phase equilibrium is approached (Gombosi 1994). Therefore, pharyngeal collapse should allow for the
condensation of more vapor from exhaled air before it is expelled. This process will also contribute to warming as
respiratory airway vapor retention by means of condensation is exothermic (Cole 1954; McFadden et al. 1982).
The Circulatory Benefits of Nervous Stimulation
As an individual snores tissues of the tongue, the pharyngeal walls, the uvula and the tonsils collide rapidly, and
sometimes quite powerfully (Deegan et al. 1995). The direct mechanical effect of pressure and movement caused
by snoring should effectively stimulate local nerve receptors. When nerve receptors within a tissue are stimulated
sufficiently they cause the blood vessels within the proximate tissues to dilate, increasing warmth and circulation in
the surrounding area. Once stimulated by the process of snoring, nerve receptors along the upper airway will
cause surrounding blood vessels to dilate, effectively increasing circulation. Increased blood flow to the pharynx
should help to provide warmth to inhaled air, and thus to the alveoli as well.
The Circulatory Benefits of Negative Intrathoracic Pressure
The obstruction of the pharyngeal airway can create relatively large amounts of negative intrathoracic pressure
(Rollheim et al. 1999). This negative pressure creates resistance against the inhalation of air forcing the muscles
involved in respiration to work harder in order to obtain the necessary amounts of oxygen (Henke et al. 1991).
Increases in the work level and energy expenditure of these muscles should result in increased overall circulation
and warmth.
Phylogenetic Considerations
To understand the structure most significant in the process of pharyngeal obstruction, the soft palate, we should
look to its phylogenetic origins. Researchers believe that upon divergence with reptiles, mammals developed the
soft, and hard palates. The palates are thought to separate the nasal cavity from the oral cavity, enabling
mammals to breathe during food consumption (Thomason et al. 1986). A closer look into the divergence of the
reptilian and mammalian classes may provide evidence for why pharyngeal collapse presents in mammals
exclusively.
For millions of years reptiles were members of one of the most dominant classes on the earth, and yet they were,
and still are, reliant on energy from their external environment to obtain the warmth necessary for physical
activity. Around 180 million years ago warm-blooded mammals began to diverge from their ectothermic, reptilian
ancestors. Mammals adapted a way to maintain a constant body temperature through biological processes, but
this increased functionality came at a price. Mammals necessitate more energy than do reptiles in order to remain
active and they are more susceptible to relatively extreme temperatures. This increased susceptibility may have
necessitated that mammals adopt a protective homeostatic mechanism, an explicitly exaptive enlargement of the
soft palate.
Mammals seem to be the only animals susceptible to nocturnal pharyngeal collapse and the preponderance of
mammals affected by it is reflected within the literature. Collapse can affect dogs (Veasey 1999; Petrof et al.
1994), primates (Guthrie et al. 1980), seals (Milsom et al. 1996), mice (Nakamura et al. 2003), pigs (Lonergan
1998) and many other mammals as well. There is no documented evidence of the existence of either collapse or
of conspicuously enlarged pharyngeal structures in reptiles.
Unlike reptiles, mammals necessitate prolonged periods of deep sleep defined by slow wave activity (SWA) and
rapid eye movement (REM) (Siegel 1999). Insensitivity to decreasing body temperature during non-conscious,
deep sleep might have been another factor that caused mammals to be selected for more massive pharyngeal
structures. Finally, it is well established that significant reductions in pharyngeal dilation accompany rapid-eye-
movement (REM) sleep (Wiegand et al. 1991).
The Prevalence of Pharyngeal Collapse in Pregnant Mothers
It is well accepted that the sickness and vomiting associated with food poisoning act as part of an immunodefence
mechanism, protecting the human body from microbial infection. A research biologist by the name of Margie
Prophet (1995) has promulgated that the morning sickness experienced by pregnant mothers is also protective.
Prophet was able to show that mothers that are less prone to morning sickness are also statistically more likely to
have a miscarriage. She also gathered evidence to support the claim that drugs that reduce morning sickness
may be associated with fetal abnormality. Because of her work, there is now a consensus among immunologists
that the morning sickness displayed by pregnant mothers may well be an evolutionary adaptation that responds to
the susceptibility of the fetus to food poisoning.
It is well accepted that pharyngeal obstruction is more prevalent in pregnant women (Hastie et al. 1989; Lefcourt
et al. 1996), and that pregnant women report frequent snoring more often than non-pregnant women (Loube et al.
1996). Interestingly, a large amount of data has shown that the increase in the incidence of OSAS in pregnant
mothers is very slight (Brownell et al. 1986; Hertz et al. 1992), that pregnancy is not generally associated with the
precipitation of OSAS in non-obese women (Maasilta et al. 2001) and that self reported somnolence does not
seem to increase in pregnancy as might be expected (Brownell et al. 1986; Hertz el al. 1982). Furthermore,
mothers that experienced obstructive sleep apnea during pregnancy did not seem to be at any significant risk for
delivering infants with fetal compromise (Loube et al. 1996). At first these findings seem anomalous but a closer
look provides clarity.
The mechanism for increased upper airway resistance in pregnancy is diffuse edema (the accumulation of fluid)
within the pharynx (Holdcroft et al. 1977). This edema is very consistent in pregnant women (Schnider et al.
1977), as is the mechanism that mitigates it. Increased pharyngeal muscle tone, possibly caused by elevated
levels of progesterone, is thought to be the predominant factor involved in helping pregnant women overcome
apnea by compensating for pharyngeal edema (Popovic et al. 1995; Loube et al. 1996).
Drawing conclusions similar to those drawn by Margie Prophet, it appears that pregnant mothers may have
evolved a protective increase in pharyngeal obstruction and snoring, but also have evolved a way to compensate
for the negative effect of obstruction, apnea. Even if sleep apnea were to result in depleted oxygen saturation in
a sleeping mother, there is reason to believe that her fetus might remain relatively unharmed because of certain
properties particular to fetal circulation and the fetal hemoglobin (Awe et al. 1979). The current article contends
that like morning sickness, pharyngeal collapse is protective, and that an increase in the incidence and severity of
collapse in pregnant mothers may allow them to better regulate homeostasis, and better protect the fetus.
Results
We have sufficiently satisfied some of the criteria that Williams and Nesse suggest should be required for all cases
of evolutionary medicine. For example, we have shown that pharyngeal collapse is relatively prevalent (Bearpark
et al. 1995; Ohayon et al. 1997; Young et al. 1993), that it is heritable (Davies et al. 1996; Silverberg et al. 1997)
and that vulnerability varies in both human and animal populations (Gau et al. 1995; Guthrie et al. 1980; Hui et al.
1999; Redline et al. 2002). We have also shown that the sequelae associated with pharyngeal collapse may well
lower reproductive success by decreasing health and lowering life expectancy (Shin et al. 2003). Despite the
possibility that the symptoms strongly associated with pharyngeal collapse, including hypoxia, hypoxemia, cluster
headaches, somnolence, fetal abnormality, brain damage and death, created evolutionary disadvantages for our
phylogenetic ancestors, the causal, physiological structures remain prevalent throughout present day human and
mammalian populations.
We have attempted to explain this anomaly, and satisfy the last criterion proposed by Williams and Nesse, by
hypothesizing that certain homeostatic properties that arise with collapse provide adaptive benefits that outweigh
the associated sequelae. Pharyngeal collapse very appropriately arises exclusively during sleep, a period that
renders many animals vulnerable to heat loss. By decreasing the distance between pharyngeal walls the
constriction of the pharynx allows for more intermolecular collisions within inspired air. These collisions should
help to warm and humidify inspired air before it reaches the alveoli, reproducing the benefits afforded to mammals
by reflex bronchoconstriction. We have also discussed how pharyngeal collapse may enable increased circulation
and warmth within the thoracic cavity by inducing increased muscular activity and by creating mechanical
stimulation.
We have also mentioned that methods of surgical intervention have proven that pharyngeal obstruction and the
sleep related disorders that accompany it can be effectively ameliorated by the reduction of certain fleshy
structures near the pharynx. The reduction of these structures poses no threat to patients and thus it is not clear
what function their excessive size may serve besides those promulgated by the present article. The relative
degree of hypotonicity of the pharyngeal musculature as well as the relative size of the soft palate, the uvula and
the pharyngeal walls may well be protective and in modern times, vestigial.
Conclusions
The concept of evolutionary medicine has become an increasingly emergent theme in medical science and it has
profoundly influenced the viewpoints assumed by immunologists and pathologists. It has become well accepted
that certain forms of disease can be precipitated by specific physiological or immunological adaptations which
arise to combat more pernicious diseases. For example, sickle cell anemia is an unfortunate complication of an
evolutionary response to malaria just as cystic fibrosis is a similar response to cholera. Individuals that have a
moderate genetic propensity for such adaptive responses are able to successfully combat the specific disease.
However, those that have a strong genetic propensity (often characterized by homozygosity) for such a response
can experience unfortunate, debilitating complications. In this light, it is interesting to note that the propensity for
habitual snoring and OSAS is heritable (Silverberg et al. 1997), with severe complications associated with
combined maternal and paternal proclivity (Redline et al. 2002).
Researcher John Remmers (2001) finds it “puzzling” that autonomic pharyngeal dilation, an “essential reflex,” is
reduced or lost during sleep. If pharyngeal collapse had no associated benefit, would not evolution have selected
those mammals with more robust pharyngeal musculature and smaller, obstructive pharyngeal structures?
Wouldn’t natural selection have responded to the disadvantages by excising the genotypic expressions of these
structures as precisely and effectively as present day otolaryngological surgeons do of their phenotypic
counterparts?
Considerations Related to Future Research
The hypothesis promulgated by this article cannot be adequately substantiated because of the dearth of related
research. As such, like most discussions related to macroevolution, this article has resorted to inferring process
from pattern. It is evident that much more work is needed to define the parameters of the influence of pharyngeal
collapse on homeostasis.
If airway collapse does influence out-of-doors nocturnal respiration then there should be disease states
secondary to its absence. Carefully designed studies that compare the internal and alveolar temperatures in
collapse prone animals with those of collapse resistant animals must be performed. Definitive knowledge of the
effects of pharyngeal collapse should prove useful for medical science. If collapse really does confer a significant
homeostatic benefit then it will be essential for physicians to counsel their uvulopalatopharyngoplasty patients
accordingly.
References:
Anuntaseree, W., K. Rookkapan and S. Kuasirikul et al. (2001) Snoring and obstructive sleep apnea in Thai
school-age children: prevalence and predisposing factors. Pediatr Pulmonol 32, 222-227.
Awe, R., M. Nicotra and T. Newsom (1979) Arterial oxygenation and alveolar-arterial gradients in term pregnancy.
Obstet Gynecol 53,182-186.
Barbet, J.P., M. Chauveau, S. Labbe and A. Lockhart (1988) Breathing dry air causes acute epithelial damage
and inflammation of the guinea pig trachea. J Appl Physiol 64, 1851-1857.
Bearpark, H., L. Elliott and R. Grunstein et al. (1995) Snoring and sleep apnea: a population study in Australian
men. Am J Respir Crit Care Med 151,1459-1465.
Brouillette R., S. Fernbach and C. Hunt (1982) Obstructive sleep apnea in infants and children. J Pediatr 100(1),
31-40.
Brownell, L., P. West and M. Kryger (1986) Breathing during sleep in normal pregnant women. AM Rev Respir Dis
133, 38-41.
Cirignotta, F., R. D’Alessandro and M. Partinen et al. (1989) Prevalence of every night snoring and obstructive
sleep apnoeas among 30–69-year-old men in Bologna, Italy. Acta Neurol Scand 79, 366-372.
Cole, P. (1954) Recordings of respiratory air temperature. J. Laryngol Otol 68, 295-307.
Corbo, G.M., F. Fuciarelli and A. Foresi et al. (1989) Snoring in children: association with respiratory symptoms
and passive smoking. BMJ 299,1491-1494.
Crumley, R.L., M. Stein and G. Gamsu et al. (1987) Determination of obstructive sleep apnea. AJR Am J
Roentgenol 148, 1069-1074.
Davies, R. and J. Stradling (1996) The epidemiology of sleep apnea. Thorax 51(suppl 2),S65-S70.
Deegan, P. and W. McNichols (1995) Pathophysiology of obstructive sleep apnoea. Eur Respir J 8(7), 1161-78.
Doyle, W.J., D. Skoner, J. Seroky, P. Fireman and J. Gwaltney (1994) Effect of experimental rhinovirus 39 on the
nasal response to histamine and cold air challenges in allergic and nonallergic subjects. J Allergy Clin Immunol 93,
534-42.
Eagan, C.J. (1963) Local vascular adaptations to cold in man. Federation Proc. 22, 947-951.
Ferrus, L., H. Guenard, G. Vardon and P. Varene (1980) Respiratory water loss. Respir Physiol 39, 367-381.
Fitzpatrick, M., K. Martin and E. Fossey et al. (1993) Snoring, asthma and sleep disturbance in Britain: a
community-based survey. Eur Respir J6, 531-535.
Gau, S. and W. Soong. (1995) Sleep problems of junior high school students in Taipei. Sleep 18, 667-673.
Geller, D., W. Morgan, K. Cota, A. Wright and L. Taussig (1988) Airway responsiveness to cold, dry air in normal
infants. Pediatr Pulmonol 4, 90-97.
Gombosi, T. (1994) Gaskinetic Theory. Cambridge, Cambridge University Press.
Guthrie, R., T. Standaert, W. Hodson and D. Woodrum (1980) Sleep and maturation of eucapnic ventilation and
CO2 sensitivity in the premature primate. J Appl Physiol 48, 347-354.
Hastie, S., K. Prowse and W. Perks et al. (1989) Obstructive sleep apnoea during pregnancy requiring
tracheostomy. Aust NZ J Obstet Gynaecol 3, 365-367.
Henke K., J. Dempsey, M. Badr, J. Kowitz and J. Skatrud (1991) Effect of sleep-induced increases in upper airway
resistance on respiratory muscle activity. J Appl Physiol 70, 158-168.
Hertz, G., A. Fast and S. Feinsilver et al. (1992) Sleep in normal late pregnancy. Sleep 15, 246-51.
Heym, J., G. Steinfels and B. Jacobs (1982) Activity of serotonin containing neurons in the nucleus raphe pallidus
of freely moving cats. Brain Res 251, 259-276.
Hoffstein, V. (1996) Snoring. Chest 109, 201-222
Holdcroft A., D. Bevan and E. O’Sullivan et al. (1977) Airway closure during pregnancy. Anesthesia 32, 517-523.
Horner, R., J. Innes, M. Morrell, S. Shea and A. Guz (1994) The effect of sleep on reflex genioglossus muscle
activation by stimuli of negative airway pressure in humans. J Physiol (Lond) 476,141-151.
Horner, R. (1996) Motor control of the pharyngeal musculature and implications for the pathogenesis of
obstructive sleep apnea. Sleep 19, 827-853.
Hui, D., J. Chan and A. Ho et al. (1999) Prevalence of snoring and sleep-disordered breathing in a student
population. Chest 116,1530-1536.
Iscoe S., R. Young and D. Jennings (1983) Control of respiratory pattern in conscious dog: effects of heat and
CO2. J Appl Physiol 54, 623-631.
Jennum, P. and A. Sjol (1993) Snoring, sleep apnoea and cardiovascular risk factors: the MONICA II Study. Int J
Epidemiol 23, 439-444
Kapur, V., K. Strohl, and S. Redline et al. (2002) Underdiagnosis of sleep apnea syndrome in US communities.
Sleep Breath 6, 49-54.
Krespi, Y., A. Keidar and M. Khosh et al. (1994) The efficacy of laser-assisted uvulopalatoplasty in the
management of obstructive sleep apnea and upper airway resistance syndrome. Otolaryngol Head Neck Surg 6,
235-243.
Kubin, L., H. Tojima, C. Reignier, A. Pack and R. Davies (1996) Interaction of serotonergic excitatory drive to
hypoglossal motoneurons with carbachol-induced, REM sleep-like atonia. Sleep 19, 187-195.
Lavie, P., L. Lavie and A. Perelman (2001) Plasma Homocysteine Levels in Obstructive Sleep Apnea: Association
With Cardiovascular Morbidity. Chest 120, 900-908.
Lefcourt, L. and J. Rodis (1996) Obstructive sleep apnea in pregnancy. Obstet Gynecol Survey 51, 503-506.
Liistro, G., D. Stanescu, G. Dooms, D. Rodenstein and C. Veriter (1988) Head position modifies upper airway
resistance in men. J Appl Physiol 64(3), 1285-1288.
Lloyd, E. (1994) ABC of sports medicine: temperature and performace I: cold. BMJ 309, 531-534.
Lonergan III, R., J. Ware, R. Atkinson, W. Winter and P. Suratt (1998) Sleep apnea in obese miniature pigs. J Appl
Physiol 84(2), 531-536.
Loube, D., J. Poceta and M. Morales et al. (1996) Self-reported snoring in pregnancy. Association with fetal
outcome. Chest 109, 885-889.
Lugaresi, E., F. Cirignotta and G. Coccagna et al. (1980) Some epidemiological data on snoring and
cardiocirculatory disturbances. Sleep 3, 221-224.
Maasilta, P., A. Bachour and K. Terama et al. (2001) Sleep-related disordered breathing during pregnancy in
obese women. Chest 120, 1448-1454.
Mc Fadden, J. (1983) Respiratory heat and water exchange: physiological and clinical implications. J Appl Physiol:
Respirat Environ Exercise Physiol 54(2), 331-336.
Mc Fadden, J., D. Denison, J. Waller, B. Assoufi, A. Peacock and T. Sopwith (1982) Direct recordings of the
temperatures in the tracheobronchial tree in normal man. J Clin Invest 69, 700-705.
Millar, J., J. Nairn, R. Unkles and R. Mcneil (1965) Cold air and ventilatory function. BR J Dis Chest 59, 23-27.
Milsom, W., M. Castellini, M. Harris, J. Castellini, D. Jones, R. Berger, S. Bahrma, L. Rea and D. Costa (1996)
Effects of hypoxia and hypercapnia on patterns of sleep-associated apnea in elephant seal pups. Am J Physiol
Regulatory Integrative Comp Physiol 271, R1017-1024.
Morin, D., R. Monteau and G. Hilaire (1992) Compared effects of serotonin on cervical and hypoglossal inspiratory
activities: an in vitro study on the newborn rat. J Physiol 451, 605-629.
Nakamura, A, Y. Fukuda and T. Kuwaki (2003) Sleep apnea and effect of chemostimulation on breathing
instability in mice. J Appl Physiol 94(2), 525-532.
Nesse R, and Williams G. (1998) Evolution and the origins of disease. Scientific American 279: 58-65.
O’Cain, C., N. Dowling, A. Slutsky, M. Hensley, K. Strohl, J. McFadden and J. Ingram (1980) Airway effects of
respiratory heat loss in normal subjects. J Appl Physiol: Respirat Environ Exercise Physiol 49, 875-880.
Ohayon, M., C. Guilleminault, R. Priest et al. (1997) Snoring and breathing pauses during sleep: telephone
interview survey of a United Kingdom population sample. BMJ 314, 860–863.
Petrof, B., A. Pack, A. Kelly, J. Eby and J. Hendricks (1994) Pharyngeal myopathy of loaded upper airway in dogs
with sleep apnea. J Appl Physiol 76, 1746-1752.
Popovic, R. and D. White (1995) Influence of gender on waking genioglossal electromyogram and upper airway
resistance. Am J Respir Crit Care Med 152, 725-31.
Powell, N., R. Riley, R. Troell et al. (1998) Radiofrequency volumetric tissue reduction of the palate in subjects
with sleep-disordered breathing. Chest 133, 1163-1174.
Proctor, D. and D. Swift (1977) Temperature and water vapor adjustment. In: Respiratory Defense
Mechanisms, edited by JD Brain, DF Proctor, and LM Reid. New York: Dekker, vol. 5, part 1, p. 95-125. (Lung Biol
Health Dis Ser)
Profet, M. (1995) Protecting Your Baby-To-Be. Reading, Mass, Addison Wesley.
Rappai, M., N. Collop, K. Stephen and R. deShazo (2003) The nose and sleep-disordered breathing: what we
know and what we do not know. Chest 124, 2309-2323.
Redline, S. and P. Tishler (2000) The genetics of sleep apnea. Sleep Med Rev 4, 583-602.
Redline, S., P. Tishler and M. Hans et al. (1997) Racial differences in sleep-disordered breathing in African-
Americans and Caucasians. Am J Respir Crit Care Med 155, 186-192.
Remmers, J. (2001) Wagging the tongue and guarding the airway: reflex control of the genioglossus. Am J Respir
Crit Care Med 164(11), 2013-2014.
Powell, N., R. Riley and R. Troell et al. (1998) Radiofrequency volumetric tissue reduction of the palate in subjects
with sleep-disordered breathing. Chest 113, 1163-1174.
Roehrs, T., W. Conway and R. Wittig et al. (1985) Sleep-wake disturbances in patients with sleep-related
respiratory disorders. Am Rev Respir Dis 132, 520-523
Rollheim, J., T. Osnes and H. Miljeteig (1999) The sites of obstruction in OSA, identified by continuous
measurements of airway pressure and flow during sleep: ambulatory vs in-hospital recordings. Clin Otolaryngol
24, 502-506.
Serway, R. (1990) Physics for Scientists and Engineers with Modern Physics, 3rdEd, Saunders College Publishing.
Shepard, J. Jr (1985) Gas exchange and hemodynamics during sleep. Med Clin North Am DA 19851231(6), 1243-
1264.
Shin, C., S. Joo, J. Kim and T. Kim (2003) Prevalence and Correlates of Habitual Snoring in High School Students.
Chest 124, 1709-1715.
Shnider, S and G. Levinson (1977) Anesthesia for obstetrics. In: Miller RD, ed. Anesthesia 32, 517-523.
Siegel, J. (1999) The evolution of REM sleep. In Handbook of Behavioral State Control., Lydic R and Baghdoyan
eds., pp 87-100, CRC Press, Boca Raton.
Silverberg, D. and A. Oksenberg (1997) Essential hypertension and abnormal upper airway resistance during
sleep. Sleep 20, 794-806.
Teran-Santos, J., A. Jimenez-Gomez, J. Cordero-Guevara (1999) The association between sleep apnea and the
risk of traffic accidents. N Engl J Med 340, 847-851.
Thomason, J. and A. Russell (1986) Mechanical factors in the evolution of the mammalian secondary palate: a
theoretical analysis. Morphol 189(2),199-213.
Togias, A., R. Naclerio, and S. Peters et al. (1986) Local generation of sulfidopeptide leukotrienes upon nasal
provocation with cold, dry air. Am Rev Respir Dis 133, 1133-1137.
Utley, D., E. Shin, A. Clerck et al. (1997) A cost-effective and rational surgical approach to patients with snoring,
upper airway resistance syndrome, or obstructive sleep apnea syndrome. Laryngoscope 107, 726-734.
Veasey, S., P. Fenik, K. Panckeri, A. Pack and J. Hendricks (1999) The Effects of Trazodone with L-Tryptophan
on Sleep-disordered Breathing in the English Bulldog. Am J Respir Crit Care Med 160(5), 1659-1667.
Wark, K. and D. Richards (1999) Thermodynamics, 6th Ed., McGraw-Hill.
Wiegand, L., C. Zwillich, D. Wiegand and D. White (1991) Changes in upper airway muscle activation and
ventilation during phasic REM sleep in normal men. J Appl Physiol 71, 488-497.
Williams, G. and R. Nesse (1991) The Dawn of Darwinian Medicine. The Quarterly Review of Biology 66, 1-22.
Young, T., L. Evans, L. Finn and M. Palta (1997) Estimation of the clinically diagnosed proportion of sleep apnea
syndrome in middle-aged men and women. Sleep 20, 705-706.
Young, T., M. Palta, J. Dempsey, J. Skatrud, S. Weber, S. Badr (1993) The occurrence of sleep-disordered
breathing among middle-aged adults. N Engl J Med 32, 1230-1235.
Young, T., P. Peppard, M. Palta, K. Hla, L. Finn, B. Morgan and J. Skatrud (1997) Population-based study of
sleep-disordered breathing as a risk factor for hypertension. Arch Intern Med 157, 1746-1752.
Zielinski, J., A. Zgierska, M. Polakowska et al. (1999) Snoring and excessive daytime somnolence among Polish
middle-aged adults. Eur Respir J 14, 946-950.
The Role of Pharyngeal Collapse in Nocturnal Homeostasis:
Exploring the Benefits and Costs of a Vastly Prevalent Phenomenon under an Evolutionary Framework
Jared Reser 06/05/04
Please send all inquiries to:
jared@jaredreser.com
Abstract
The present article will explore how the collapse of the pharyngeal airway during sleep, a phenomenon currently
identified as an indicator of disease, may confer specific homeostatic benefits. More specifically, we will consider
how pathologically large size of the fleshy structures in the pharyngeal airway, along with relative hypotonicity of
the pharyngeal musculature may have provided our ancestors a selective advantage by enabling them to reduce
nocturnal, respiratory heat loss. When the pharynx is collapsed, the diameter of the pharyngeal airway
decreases, allowing increased intermolecular collisions between inspired air and the epithelial walls of the upper
airway. An increase in the number of collisions should facilitate the transference of both warmth and humidity to
inspired air before it reaches the more sensitive lower airway resulting in the maintenance of internal, core
temperature and alveolar heat retention. By inducing increased muscular activity in the diaphragm and by
mechanically stimulating specific pharyngeal structures, pharyngeal collapse might provide other homeostatic
benefits as well. In light of the debilitating disorders associated with pharyngeal collapse, the absence of a
disease state secondary to surgical correction, the high proclivity for collapse documented in pregnant mothers,
and the functional similarities between collapse and reflex bronchoconstriction, it appears that the phenomenon is
explicable under evolutionary medicinal theory.
Keywords
bronchoconstriction, genetic epidemiology, hypotonia, mean free path, obstructive sleep apnea syndrome,
pharyngeal collapse, pulmonary mechanics
Introduction
The collapse of the pharyngeal airway, a physiological occurrence that accompanies sleep, is known to affect all
people. The degree of collapse experienced by an individual is determined by his or her genetic vulnerability and
can be placed along a spectrum from healthy breathing to apnea and disease. In susceptible individuals,
pharyngeal collapse presents as obstructive sleep apnea syndrome (OSAS), and upper airway resistance
syndrome (UARS), two widely prevalent, relatively pernicious disorders.
Using a paradigm constructed by Williams and Nesse (1991) this article will attempt to analyze pharyngeal
collapse using evolutionary medicinal theory. In order to show that a trait may be a form of evolutionary medicine,
under this framework, it is necessary to provide evidence that the trait is relatively prevalent, that it is heritable
and that susceptibility varies within populations. It is also necessary to show how the genes responsible for the
trait might have been naturally selected. For example, many heritable traits involve a tradeoff or compromise,
and, when analyzing these, it is necessary to provide evidence showing that the decrement in reproductive
success due to associated health risks can be mitigated by associated health benefits (Williams & Nesse 1991).
Williams and Nesse have identified many such “pathological” conditions such as anxiety, cough, diarrhea, fever,
inflammation, pain, sneezing and vomiting and have helped to show that they are actually evolved defenses
(1998).
This article will examine how pharyngeal collapse might create a net fitness advantage by playing a passive but
significant role in the maintenance of nocturnal homeostasis. Today, the costs of collapse are well documented
but the defensive manifestations may be hidden because of compensatory aspects of our modern environment
(ie. insulation and central heating). Providing an evolutionary explanation for susceptibility to collapse may help
explain the high prevalence in human populations and may inform both medical and gene therapy. Before we
consider the ultimate or evolutionary cause of pharyngeal collapse we must first look at the proximate,
pathophysiological causes. Because the present hypothesis is in part predicated on pathophysiological data, the
provision of background information describing the etiology, symptoms, and methods of intervention is in order.
Etiology of Pharyngeal Collapse
The discussion of etiology motivates us to focus on two causal factors, namely the collapse caused by hypotonia
(relaxation) in pharyngeal musculature, and the obstruction caused by specific structures within the pharynx.
Pharyngeal collapse arises when the airway that allows easy and normal breathing becomes blocked. When a
person is awake pharyngeal dilator muscles maintain a clearly defined, open pathway to the lungs. These
muscles disallow specific, membranous structures within the airway from interfering with inhalation. When a
person is asleep, however, these muscles become hypotonic; they relax and may tend to collapse (Horner 1996).
It is well accepted that the nocturnal collapse of pharyngeal musculature is responsible for both the increase in the
degree of obstruction caused by specific pharyngeal structures, and the restriction of the flow of air through the
upper and pharyngeal airways (Horner et al. 1994; Wheatley et al. 1993).
The sound associated with severe pharyngeal obstruction, snoring, occurs when certain structures within the
pharyngeal airway vibrate excessively as air is pulled through the narrow opening, toward the lungs. Any of the
membranous structures that lack cartilaginous support, including the uvula, adenoids, soft palate, pharyngeal
walls, and tonsillar pillars, are predisposed to vibration (Crumley et al. 1987). The soft palate (the upper surface
of the mouth that separates the oral and nasal cavities) and the uvula (the small piece of soft tissue that hangs
from the soft palate) are relatively large and fleshy, and can obstruct the upper airway completely. When these
structures obstruct the upper airway for even a short period of time they may cause discomfort or morbidity.
Sequelae and Associated Disorders
The complications associated with pharyngeal collapse are numerous and they vary in severity. A review of these
complications allows insight into how such a potent dysfunction might have seriously jeopardized the evolutionary
fitness of our ancestors.
Repetitive and severe obstruction of the pharyngeal airway results in the upper airway resistance syndrome
(UARS), a form of sleep disordered breathing in which resistance to airflow leads to brief arousals from sleep
thereby causing daytime somnolence (weariness) (Zielinski et al. 1999). Many UARS sufferers experience
episodes of severe obstruction lasting from a few seconds to a full minute. These episodes take two forms:
hypopnea (periods of reduced breathing) and apnea (cessation of breathing). The related disorder, obstructive
sleep apnea syndrome (OSAS), often results in a loss of oxygen which can create serious disturbances in health.
The cessation of breath caused by obstruction can induce hypoxemia, a state defined by insufficient oxygen in the
blood (oxyhemoglobin desaturation). It has been estimated that 24% of men between the ages of 30 and 60
suffer from apnea induced hypoxemia (Young et al. 1997). Painful cluster headaches precipitated by hypoxemia
are also strongly associated with OSAS (Kudrow and Kudrow 1990, 1993).
Prolonged hypoxemia often results in hypoxia, a deficiency in the amount of oxygen reaching body tissues.
Hypoxia induced by OSAS is known to cause tissue damage in various regions of the body (Jennum et al. 1993),
including the brain. It has been suggested that hypoxia caused by OSAS is linked with hypertension (Young et al.
1997), heart disease (Lugaresi et al. 1980), cerebrovascular disease (Neau et al. 1995), and various other
cardiovascular diseases (Lavie et al. 2001). Recognition of the dangers associated with OSAS within the
literature is a cause of increasing public interest and concern (Shin et al. 2003).
Medical Intervention
An analysis of the methods of medical intervention reveals that the causal pharyngeal structures can be reduced
surgically with virtually no negative consequence. There are several methods of intervention aimed at mitigating
pharyngeal obstruction and they range from oral devices to surgery. The most successful way to stop habitual
snoring is the surgical removal of portions of the soft palate (Krespi et al. 1994). Doctors that practice head and
neck surgery, otolaryngology, perform such operations not as a last resort, but as a relatively successful method
of medical intervention (Utley et al. 1997). Such surgical procedures include uvulopalatopharyngoplasty (UPPP),
Laser-Assisted Uvuloplasty (LAUP) and palatal reduction by radio frequency ablation (somnoplasty). Larger
populations of patients must be analyzed before surgery can be recommended as the primary form of therapy;
however, there are currently no persisting, negative complications associated with the surgical reduction of the
soft palate (Powell et al. 1998). Because the reduction of the soft palate poses no threat to modern day patients,
an evolutionary explanation for the wide prevalence of its excessive size should prove informative.
The Prevalence of Pharyngeal Collapse
Pharyngeal collapse is a phenomenon that affects all people and it can result in various disorders that are highly
prevalent in human populations around the world. Studies have demonstrated that the prevalence of OSAS could
vary between 3.2% and 12.1% in children (Corbo et al. 1989; Anuntaseree et al. 2001), and between 5% and 40%
in adults (Bearpark et al. 1995; Cirignotta et al. 1989; Fitzpatrick et al. 1993; Redline et al. 1997). Also elderly
populations are at significantly higher risk than younger populations (Young et al. 1993). Snoring is also very
common in the general population; 35–45% of men and 15–28% of women report that they snore habitually
(Ohayon et al. 1997; Young et al. 1993). The elderly, adults, children and even infants (Brouillette et al. 1982)
experience pharyngeal collapse and the associated sequelae. The presence of collapse throughout the
maturational timeline is indicative of its value as a protective mechanism.
Like most inherited traits, and protective adaptations, a person’s proclivity for pharyngeal collapse is genetic
(Davies et al. 1996; Silverberg et al. 1997). Many researchers consider it likely that inherited, familial factors such
as upper airway structure and neural control of the upper airway muscles interact to produce relative proclivity for
OSAS (Redline et al. 2002). Because environmental factors can vary greatly from area to area, the relative
distribution of most protective traits is known to vary widely, even between members of the same species. As
might be expected, a large amount of variance exists with regard to the severity of pharyngeal collapse within the
human species. Wide ethnic variations (Redline et al. 1997) in the prevalence of habitual snoring might result from
differences in definitions and methodology, but more importantly it may stem from differences in population
genetics. In fact both the incidence of habitual snoring and the number of studies describing OSAS appear to be
far less prevalent in Asia (Gau et al. 1995; Hui et al. 1999).
Hypothesis: Pharyngeal Collapse May Reduce Respiratory Heat Loss
If the risks associated with pharyngeal collapse make it a potentially enervating and morbid activity, how then
could it have become so ubiquitous? Why would the human body so dramatically decrease autonomic, muscular
support to the throat during rest, and why would such large, fleshy structures exist in an area so vital to respiration
and health? Despite the scarcity of related research the present article will attempt to characterize pharyngeal
collapse as a mechanism, much like other respiratory mechanisms, that promotes the maintenance of
homeostasis by regulating respiratory heat retention. Collapse occurs exclusively during sleep- this may be a
functional response to the cold exposure that accompanies sleep related processes.
Factors that Expose Sleeping Animals to the Cold
There are at least two factors that expose diurnal, sleeping animals to the cold. When an animal is sleeping its
skeletal muscles are quiescent, especially during deep sleep. A lack of muscular exertion is a common cause of
reduction in internal temperature. Also, it is widely recognized that the temperature of nighttime air is relatively low
compared to daytime air. This temperature difference exposes diurnal animals to the cold when they are the most
vulnerable. For these two reasons diurnal animals are particularly susceptible to heat loss during sleep.
The sleeping mammal’s respiratory tract is a prime candidate for the cooling process because it contains many
square inches of wet, soft tissue that is exposed to rapidly moving, dry air for many hours at a time (Eagan 1963).
As each new breath of air is inhaled, water molecules evaporate from the throat and lungs as part of an
endothermic process, and a potentially large amount of body heat can be lost (Ferrus et al. 1980). For example,
dogs utilize the respiratory tract as their primary means of dissipating excessive heat (Iscoe et al. 1983).
Humans are susceptible to substantial changes in external temperatures and will die if their core, internal
temperature drifts more than a few degrees above or below 37° C (Lloyd 1994). It is fortunate that humans, as
homeothermic mammals, are thermoregulators. They utilize specific homeostatic mechanisms to minimize the heat
lost to the external environment. Such mechanisms include vasoconstriction, concurrent heat exchange,
metabolic regulation and shivering. These methods of heat conservation focus on maintaining the core, internal
temperature at the expense of heat loss within the extremities. Heat loss arising from respiration therefore can be
very potent because it conducts heat away from the lungs and thorax, the very center of the body. If it is true that
nocturnal, sleeping processes are more susceptible to heat loss than diurnal, wakeful processes, and it is true
that pharyngeal collapse is a mechanism involved in minimizing the rate of heat exchange between the respiratory
system and the surrounding environment, then the involvement of collapse as an exclusively nocturnal mechanism
is quite appropriate.
The Importance of Heating Inspired Air
The respiratory airways both heat and humidify inspired air before it reaches the alveoli, the final destination in
the respiratory tract (Millar et al. 1965). Researchers do not seem to have reached a consensus regarding the
benefits of maintaining the temperature of the lower airways and alveoli, but they do seem to agree on its
importance. McFadden reported that the inhalation of dry, cold air increases cardiovascular stress and
decreases pulmonary function (1983). Furthermore, certain reflexive measures taken by the respiratory system
when it is exposed to cold air have led researchers to believe that cold air is deleterious to respiratory health and
functionality (McFadden 1983). The exposure of the airways to cold air is consistently accompanied by reflex
bronchoconstriction, vasoconstriction and the release of histamines (Doyle et al. 1994; Geller et al. 1988). These
responses are regarded by researchers to be protective (O’Cain et al. 1980).
Contact of the skin with cold air is well known to stimulate thermally sensitive receptors that produce reflexive
bronchoconstriction (Doyle et al. 1994). The constriction of the passageways within the tracheo-bronchial tree
(the branches leading to the alveoli) in response to cold air is a process that diminishes susceptibility to heat and
humidity loss (O’Cain et al. 1980). In other words, the heat and humidity retention that I am ascribing to the
collapse of the pharyngeal airway has been previously ascribed to the collapse of the tracheo-bronchial tree; the
two mechanisms are physically and functionally analogous. The benefits conferred by reflex bronchoconstricion
can arise when the animal is both asleep and awake. Unlike reflex bronchoconstriction, pharyngeal collapse
responds specifically to the heat loss associated with nocturnal processes. A careful, physical analysis shows that
collapse should increase respiratory heat conservation by increasing the rate of molecular collisions within the
pharyngeal airway.
The Collapse of the Pharynx Should Result in the Warming of Inspired Air
Pharyngeal collapse should help to warm inhaled air before it reaches the lungs by creating a “narrower tube” for
air to travel through. This narrow tube should also provide other benefits including the retention of water vapor,
and the filtration of inspired pathogens. It should seem conceivable that a broader airway should not prove to be
as effective as a narrower airway in achieving these benefits. For clarification, let us turn to an academically
popular, thermodynamic system that is structurally analogous to the collapsible, pharyngeal airway.
From a physical perspective a “narrowing airway” is a cylinder that exhibits a variance in the distance between its
walls. The number of times that a gas molecule within such a cylinder will collide with the walls of its container is a
function of the distance between the walls (Wark et al. 1999). As the walls draw nearer, the average distance that
the molecules will travel before they collide with one of the walls decreases. In other words, a reduction in the
distance between walls results in an increase in the number of collisions per time period (Serway 1990).
The number of collisions is very important because it determines how much kinetic energy will be transferred from
the walls to the air molecules by conduction (Serway 1990). Knowledge of the corollaries of this physical model
allows us to conclude that a narrower airway will create more collisions between the cold air molecules and the
warm epithelial tissues of the pharyngeal airway thereby increasing the temperature of inspired air before it
reaches the alveoli.
The Conservation of Water Vapor
Considerable water loss takes place through exhalation (Ferrus et al. 1980), and by narrowing the airway, more
water vapor should be conserved. As mentioned before, a decrease in the distance between pharyngeal walls
produces more molecule-wall collisions. The kinetic theory of gasses predicts that as the number of collisions
increases, phase equilibrium is approached (Gombosi 1994). Therefore, pharyngeal collapse should allow for the
condensation of more vapor from exhaled air before it is expelled. This process will also contribute to warming as
respiratory airway vapor retention by means of condensation is exothermic (Cole 1954; McFadden et al. 1982).
The Circulatory Benefits of Nervous Stimulation
As an individual snores tissues of the tongue, the pharyngeal walls, the uvula and the tonsils collide rapidly, and
sometimes quite powerfully (Deegan et al. 1995). The direct mechanical effect of pressure and movement caused
by snoring should effectively stimulate local nerve receptors. When nerve receptors within a tissue are stimulated
sufficiently they cause the blood vessels within the proximate tissues to dilate, increasing warmth and circulation in
the surrounding area. Once stimulated by the process of snoring, nerve receptors along the upper airway will
cause surrounding blood vessels to dilate, effectively increasing circulation. Increased blood flow to the pharynx
should help to provide warmth to inhaled air, and thus to the alveoli as well.
The Circulatory Benefits of Negative Intrathoracic Pressure
The obstruction of the pharyngeal airway can create relatively large amounts of negative intrathoracic pressure
(Rollheim et al. 1999). This negative pressure creates resistance against the inhalation of air forcing the muscles
involved in respiration to work harder in order to obtain the necessary amounts of oxygen (Henke et al. 1991).
Increases in the work level and energy expenditure of these muscles should result in increased overall circulation
and warmth.
Phylogenetic Considerations
To understand the structure most significant in the process of pharyngeal obstruction, the soft palate, we should
look to its phylogenetic origins. Researchers believe that upon divergence with reptiles, mammals developed the
soft, and hard palates. The palates are thought to separate the nasal cavity from the oral cavity, enabling
mammals to breathe during food consumption (Thomason et al. 1986). A closer look into the divergence of the
reptilian and mammalian classes may provide evidence for why pharyngeal collapse presents in mammals
exclusively.
For millions of years reptiles were members of one of the most dominant classes on the earth, and yet they were,
and still are, reliant on energy from their external environment to obtain the warmth necessary for physical
activity. Around 180 million years ago warm-blooded mammals began to diverge from their ectothermic, reptilian
ancestors. Mammals adapted a way to maintain a constant body temperature through biological processes, but
this increased functionality came at a price. Mammals necessitate more energy than do reptiles in order to remain
active and they are more susceptible to relatively extreme temperatures. This increased susceptibility may have
necessitated that mammals adopt a protective homeostatic mechanism, an explicitly exaptive enlargement of the
soft palate.
Mammals seem to be the only animals susceptible to nocturnal pharyngeal collapse and the preponderance of
mammals affected by it is reflected within the literature. Collapse can affect dogs (Veasey 1999; Petrof et al.
1994), primates (Guthrie et al. 1980), seals (Milsom et al. 1996), mice (Nakamura et al. 2003), pigs (Lonergan
1998) and many other mammals as well. There is no documented evidence of the existence of either collapse or
of conspicuously enlarged pharyngeal structures in reptiles.
Unlike reptiles, mammals necessitate prolonged periods of deep sleep defined by slow wave activity (SWA) and
rapid eye movement (REM) (Siegel 1999). Insensitivity to decreasing body temperature during non-conscious,
deep sleep might have been another factor that caused mammals to be selected for more massive pharyngeal
structures. Finally, it is well established that significant reductions in pharyngeal dilation accompany rapid-eye-
movement (REM) sleep (Wiegand et al. 1991).
The Prevalence of Pharyngeal Collapse in Pregnant Mothers
It is well accepted that the sickness and vomiting associated with food poisoning act as part of an immunodefence
mechanism, protecting the human body from microbial infection. A research biologist by the name of Margie
Prophet (1995) has promulgated that the morning sickness experienced by pregnant mothers is also protective.
Prophet was able to show that mothers that are less prone to morning sickness are also statistically more likely to
have a miscarriage. She also gathered evidence to support the claim that drugs that reduce morning sickness
may be associated with fetal abnormality. Because of her work, there is now a consensus among immunologists
that the morning sickness displayed by pregnant mothers may well be an evolutionary adaptation that responds to
the susceptibility of the fetus to food poisoning.
It is well accepted that pharyngeal obstruction is more prevalent in pregnant women (Hastie et al. 1989; Lefcourt
et al. 1996), and that pregnant women report frequent snoring more often than non-pregnant women (Loube et al.
1996). Interestingly, a large amount of data has shown that the increase in the incidence of OSAS in pregnant
mothers is very slight (Brownell et al. 1986; Hertz et al. 1992), that pregnancy is not generally associated with the
precipitation of OSAS in non-obese women (Maasilta et al. 2001) and that self reported somnolence does not
seem to increase in pregnancy as might be expected (Brownell et al. 1986; Hertz el al. 1982). Furthermore,
mothers that experienced obstructive sleep apnea during pregnancy did not seem to be at any significant risk for
delivering infants with fetal compromise (Loube et al. 1996). At first these findings seem anomalous but a closer
look provides clarity.
The mechanism for increased upper airway resistance in pregnancy is diffuse edema (the accumulation of fluid)
within the pharynx (Holdcroft et al. 1977). This edema is very consistent in pregnant women (Schnider et al.
1977), as is the mechanism that mitigates it. Increased pharyngeal muscle tone, possibly caused by elevated
levels of progesterone, is thought to be the predominant factor involved in helping pregnant women overcome
apnea by compensating for pharyngeal edema (Popovic et al. 1995; Loube et al. 1996).
Drawing conclusions similar to those drawn by Margie Prophet, it appears that pregnant mothers may have
evolved a protective increase in pharyngeal obstruction and snoring, but also have evolved a way to compensate
for the negative effect of obstruction, apnea. Even if sleep apnea were to result in depleted oxygen saturation in
a sleeping mother, there is reason to believe that her fetus might remain relatively unharmed because of certain
properties particular to fetal circulation and the fetal hemoglobin (Awe et al. 1979). The current article contends
that like morning sickness, pharyngeal collapse is protective, and that an increase in the incidence and severity of
collapse in pregnant mothers may allow them to better regulate homeostasis, and better protect the fetus.
Results
We have sufficiently satisfied some of the criteria that Williams and Nesse suggest should be required for all cases
of evolutionary medicine. For example, we have shown that pharyngeal collapse is relatively prevalent (Bearpark
et al. 1995; Ohayon et al. 1997; Young et al. 1993), that it is heritable (Davies et al. 1996; Silverberg et al. 1997)
and that vulnerability varies in both human and animal populations (Gau et al. 1995; Guthrie et al. 1980; Hui et al.
1999; Redline et al. 2002). We have also shown that the sequelae associated with pharyngeal collapse may well
lower reproductive success by decreasing health and lowering life expectancy (Shin et al. 2003). Despite the
possibility that the symptoms strongly associated with pharyngeal collapse, including hypoxia, hypoxemia, cluster
headaches, somnolence, fetal abnormality, brain damage and death, created evolutionary disadvantages for our
phylogenetic ancestors, the causal, physiological structures remain prevalent throughout present day human and
mammalian populations.
We have attempted to explain this anomaly, and satisfy the last criterion proposed by Williams and Nesse, by
hypothesizing that certain homeostatic properties that arise with collapse provide adaptive benefits that outweigh
the associated sequelae. Pharyngeal collapse very appropriately arises exclusively during sleep, a period that
renders many animals vulnerable to heat loss. By decreasing the distance between pharyngeal walls the
constriction of the pharynx allows for more intermolecular collisions within inspired air. These collisions should
help to warm and humidify inspired air before it reaches the alveoli, reproducing the benefits afforded to mammals
by reflex bronchoconstriction. We have also discussed how pharyngeal collapse may enable increased circulation
and warmth within the thoracic cavity by inducing increased muscular activity and by creating mechanical
stimulation.
We have also mentioned that methods of surgical intervention have proven that pharyngeal obstruction and the
sleep related disorders that accompany it can be effectively ameliorated by the reduction of certain fleshy
structures near the pharynx. The reduction of these structures poses no threat to patients and thus it is not clear
what function their excessive size may serve besides those promulgated by the present article. The relative
degree of hypotonicity of the pharyngeal musculature as well as the relative size of the soft palate, the uvula and
the pharyngeal walls may well be protective and in modern times, vestigial.
Conclusions
The concept of evolutionary medicine has become an increasingly emergent theme in medical science and it has
profoundly influenced the viewpoints assumed by immunologists and pathologists. It has become well accepted
that certain forms of disease can be precipitated by specific physiological or immunological adaptations which
arise to combat more pernicious diseases. For example, sickle cell anemia is an unfortunate complication of an
evolutionary response to malaria just as cystic fibrosis is a similar response to cholera. Individuals that have a
moderate genetic propensity for such adaptive responses are able to successfully combat the specific disease.
However, those that have a strong genetic propensity (often characterized by homozygosity) for such a response
can experience unfortunate, debilitating complications. In this light, it is interesting to note that the propensity for
habitual snoring and OSAS is heritable (Silverberg et al. 1997), with severe complications associated with
combined maternal and paternal proclivity (Redline et al. 2002).
Researcher John Remmers (2001) finds it “puzzling” that autonomic pharyngeal dilation, an “essential reflex,” is
reduced or lost during sleep. If pharyngeal collapse had no associated benefit, would not evolution have selected
those mammals with more robust pharyngeal musculature and smaller, obstructive pharyngeal structures?
Wouldn’t natural selection have responded to the disadvantages by excising the genotypic expressions of these
structures as precisely and effectively as present day otolaryngological surgeons do of their phenotypic
counterparts?
Considerations Related to Future Research
The hypothesis promulgated by this article cannot be adequately substantiated because of the dearth of related
research. As such, like most discussions related to macroevolution, this article has resorted to inferring process
from pattern. It is evident that much more work is needed to define the parameters of the influence of pharyngeal
collapse on homeostasis.
If airway collapse does influence out-of-doors nocturnal respiration then there should be disease states
secondary to its absence. Carefully designed studies that compare the internal and alveolar temperatures in
collapse prone animals with those of collapse resistant animals must be performed. Definitive knowledge of the
effects of pharyngeal collapse should prove useful for medical science. If collapse really does confer a significant
homeostatic benefit then it will be essential for physicians to counsel their uvulopalatopharyngoplasty patients
accordingly.
References:
Anuntaseree, W., K. Rookkapan and S. Kuasirikul et al. (2001) Snoring and obstructive sleep apnea in Thai
school-age children: prevalence and predisposing factors. Pediatr Pulmonol 32, 222-227.
Awe, R., M. Nicotra and T. Newsom (1979) Arterial oxygenation and alveolar-arterial gradients in term pregnancy.
Obstet Gynecol 53,182-186.
Barbet, J.P., M. Chauveau, S. Labbe and A. Lockhart (1988) Breathing dry air causes acute epithelial damage
and inflammation of the guinea pig trachea. J Appl Physiol 64, 1851-1857.
Bearpark, H., L. Elliott and R. Grunstein et al. (1995) Snoring and sleep apnea: a population study in Australian
men. Am J Respir Crit Care Med 151,1459-1465.
Brouillette R., S. Fernbach and C. Hunt (1982) Obstructive sleep apnea in infants and children. J Pediatr 100(1),
31-40.
Brownell, L., P. West and M. Kryger (1986) Breathing during sleep in normal pregnant women. AM Rev Respir Dis
133, 38-41.
Cirignotta, F., R. D’Alessandro and M. Partinen et al. (1989) Prevalence of every night snoring and obstructive
sleep apnoeas among 30–69-year-old men in Bologna, Italy. Acta Neurol Scand 79, 366-372.
Cole, P. (1954) Recordings of respiratory air temperature. J. Laryngol Otol 68, 295-307.
Corbo, G.M., F. Fuciarelli and A. Foresi et al. (1989) Snoring in children: association with respiratory symptoms
and passive smoking. BMJ 299,1491-1494.
Crumley, R.L., M. Stein and G. Gamsu et al. (1987) Determination of obstructive sleep apnea. AJR Am J
Roentgenol 148, 1069-1074.
Davies, R. and J. Stradling (1996) The epidemiology of sleep apnea. Thorax 51(suppl 2),S65-S70.
Deegan, P. and W. McNichols (1995) Pathophysiology of obstructive sleep apnoea. Eur Respir J 8(7), 1161-78.
Doyle, W.J., D. Skoner, J. Seroky, P. Fireman and J. Gwaltney (1994) Effect of experimental rhinovirus 39 on the
nasal response to histamine and cold air challenges in allergic and nonallergic subjects. J Allergy Clin Immunol 93,
534-42.
Eagan, C.J. (1963) Local vascular adaptations to cold in man. Federation Proc. 22, 947-951.
Ferrus, L., H. Guenard, G. Vardon and P. Varene (1980) Respiratory water loss. Respir Physiol 39, 367-381.
Fitzpatrick, M., K. Martin and E. Fossey et al. (1993) Snoring, asthma and sleep disturbance in Britain: a
community-based survey. Eur Respir J6, 531-535.
Gau, S. and W. Soong. (1995) Sleep problems of junior high school students in Taipei. Sleep 18, 667-673.
Geller, D., W. Morgan, K. Cota, A. Wright and L. Taussig (1988) Airway responsiveness to cold, dry air in normal
infants. Pediatr Pulmonol 4, 90-97.
Gombosi, T. (1994) Gaskinetic Theory. Cambridge, Cambridge University Press.
Guthrie, R., T. Standaert, W. Hodson and D. Woodrum (1980) Sleep and maturation of eucapnic ventilation and
CO2 sensitivity in the premature primate. J Appl Physiol 48, 347-354.
Hastie, S., K. Prowse and W. Perks et al. (1989) Obstructive sleep apnoea during pregnancy requiring
tracheostomy. Aust NZ J Obstet Gynaecol 3, 365-367.
Henke K., J. Dempsey, M. Badr, J. Kowitz and J. Skatrud (1991) Effect of sleep-induced increases in upper airway
resistance on respiratory muscle activity. J Appl Physiol 70, 158-168.
Hertz, G., A. Fast and S. Feinsilver et al. (1992) Sleep in normal late pregnancy. Sleep 15, 246-51.
Heym, J., G. Steinfels and B. Jacobs (1982) Activity of serotonin containing neurons in the nucleus raphe pallidus
of freely moving cats. Brain Res 251, 259-276.
Hoffstein, V. (1996) Snoring. Chest 109, 201-222
Holdcroft A., D. Bevan and E. O’Sullivan et al. (1977) Airway closure during pregnancy. Anesthesia 32, 517-523.
Horner, R., J. Innes, M. Morrell, S. Shea and A. Guz (1994) The effect of sleep on reflex genioglossus muscle
activation by stimuli of negative airway pressure in humans. J Physiol (Lond) 476,141-151.
Horner, R. (1996) Motor control of the pharyngeal musculature and implications for the pathogenesis of
obstructive sleep apnea. Sleep 19, 827-853.
Hui, D., J. Chan and A. Ho et al. (1999) Prevalence of snoring and sleep-disordered breathing in a student
population. Chest 116,1530-1536.
Iscoe S., R. Young and D. Jennings (1983) Control of respiratory pattern in conscious dog: effects of heat and
CO2. J Appl Physiol 54, 623-631.
Jennum, P. and A. Sjol (1993) Snoring, sleep apnoea and cardiovascular risk factors: the MONICA II Study. Int J
Epidemiol 23, 439-444
Kapur, V., K. Strohl, and S. Redline et al. (2002) Underdiagnosis of sleep apnea syndrome in US communities.
Sleep Breath 6, 49-54.
Krespi, Y., A. Keidar and M. Khosh et al. (1994) The efficacy of laser-assisted uvulopalatoplasty in the
management of obstructive sleep apnea and upper airway resistance syndrome. Otolaryngol Head Neck Surg 6,
235-243.
Kubin, L., H. Tojima, C. Reignier, A. Pack and R. Davies (1996) Interaction of serotonergic excitatory drive to
hypoglossal motoneurons with carbachol-induced, REM sleep-like atonia. Sleep 19, 187-195.
Lavie, P., L. Lavie and A. Perelman (2001) Plasma Homocysteine Levels in Obstructive Sleep Apnea: Association
With Cardiovascular Morbidity. Chest 120, 900-908.
Lefcourt, L. and J. Rodis (1996) Obstructive sleep apnea in pregnancy. Obstet Gynecol Survey 51, 503-506.
Liistro, G., D. Stanescu, G. Dooms, D. Rodenstein and C. Veriter (1988) Head position modifies upper airway
resistance in men. J Appl Physiol 64(3), 1285-1288.
Lloyd, E. (1994) ABC of sports medicine: temperature and performace I: cold. BMJ 309, 531-534.
Lonergan III, R., J. Ware, R. Atkinson, W. Winter and P. Suratt (1998) Sleep apnea in obese miniature pigs. J Appl
Physiol 84(2), 531-536.
Loube, D., J. Poceta and M. Morales et al. (1996) Self-reported snoring in pregnancy. Association with fetal
outcome. Chest 109, 885-889.
Lugaresi, E., F. Cirignotta and G. Coccagna et al. (1980) Some epidemiological data on snoring and
cardiocirculatory disturbances. Sleep 3, 221-224.
Maasilta, P., A. Bachour and K. Terama et al. (2001) Sleep-related disordered breathing during pregnancy in
obese women. Chest 120, 1448-1454.
Mc Fadden, J. (1983) Respiratory heat and water exchange: physiological and clinical implications. J Appl Physiol:
Respirat Environ Exercise Physiol 54(2), 331-336.
Mc Fadden, J., D. Denison, J. Waller, B. Assoufi, A. Peacock and T. Sopwith (1982) Direct recordings of the
temperatures in the tracheobronchial tree in normal man. J Clin Invest 69, 700-705.
Millar, J., J. Nairn, R. Unkles and R. Mcneil (1965) Cold air and ventilatory function. BR J Dis Chest 59, 23-27.
Milsom, W., M. Castellini, M. Harris, J. Castellini, D. Jones, R. Berger, S. Bahrma, L. Rea and D. Costa (1996)
Effects of hypoxia and hypercapnia on patterns of sleep-associated apnea in elephant seal pups. Am J Physiol
Regulatory Integrative Comp Physiol 271, R1017-1024.
Morin, D., R. Monteau and G. Hilaire (1992) Compared effects of serotonin on cervical and hypoglossal inspiratory
activities: an in vitro study on the newborn rat. J Physiol 451, 605-629.
Nakamura, A, Y. Fukuda and T. Kuwaki (2003) Sleep apnea and effect of chemostimulation on breathing
instability in mice. J Appl Physiol 94(2), 525-532.
Nesse R, and Williams G. (1998) Evolution and the origins of disease. Scientific American 279: 58-65.
O’Cain, C., N. Dowling, A. Slutsky, M. Hensley, K. Strohl, J. McFadden and J. Ingram (1980) Airway effects of
respiratory heat loss in normal subjects. J Appl Physiol: Respirat Environ Exercise Physiol 49, 875-880.
Ohayon, M., C. Guilleminault, R. Priest et al. (1997) Snoring and breathing pauses during sleep: telephone
interview survey of a United Kingdom population sample. BMJ 314, 860–863.
Petrof, B., A. Pack, A. Kelly, J. Eby and J. Hendricks (1994) Pharyngeal myopathy of loaded upper airway in dogs
with sleep apnea. J Appl Physiol 76, 1746-1752.
Popovic, R. and D. White (1995) Influence of gender on waking genioglossal electromyogram and upper airway
resistance. Am J Respir Crit Care Med 152, 725-31.
Powell, N., R. Riley, R. Troell et al. (1998) Radiofrequency volumetric tissue reduction of the palate in subjects
with sleep-disordered breathing. Chest 133, 1163-1174.
Proctor, D. and D. Swift (1977) Temperature and water vapor adjustment. In: Respiratory Defense
Mechanisms, edited by JD Brain, DF Proctor, and LM Reid. New York: Dekker, vol. 5, part 1, p. 95-125. (Lung Biol
Health Dis Ser)
Profet, M. (1995) Protecting Your Baby-To-Be. Reading, Mass, Addison Wesley.
Rappai, M., N. Collop, K. Stephen and R. deShazo (2003) The nose and sleep-disordered breathing: what we
know and what we do not know. Chest 124, 2309-2323.
Redline, S. and P. Tishler (2000) The genetics of sleep apnea. Sleep Med Rev 4, 583-602.
Redline, S., P. Tishler and M. Hans et al. (1997) Racial differences in sleep-disordered breathing in African-
Americans and Caucasians. Am J Respir Crit Care Med 155, 186-192.
Remmers, J. (2001) Wagging the tongue and guarding the airway: reflex control of the genioglossus. Am J Respir
Crit Care Med 164(11), 2013-2014.
Powell, N., R. Riley and R. Troell et al. (1998) Radiofrequency volumetric tissue reduction of the palate in subjects
with sleep-disordered breathing. Chest 113, 1163-1174.
Roehrs, T., W. Conway and R. Wittig et al. (1985) Sleep-wake disturbances in patients with sleep-related
respiratory disorders. Am Rev Respir Dis 132, 520-523
Rollheim, J., T. Osnes and H. Miljeteig (1999) The sites of obstruction in OSA, identified by continuous
measurements of airway pressure and flow during sleep: ambulatory vs in-hospital recordings. Clin Otolaryngol
24, 502-506.
Serway, R. (1990) Physics for Scientists and Engineers with Modern Physics, 3rdEd, Saunders College Publishing.
Shepard, J. Jr (1985) Gas exchange and hemodynamics during sleep. Med Clin North Am DA 19851231(6), 1243-
1264.
Shin, C., S. Joo, J. Kim and T. Kim (2003) Prevalence and Correlates of Habitual Snoring in High School Students.
Chest 124, 1709-1715.
Shnider, S and G. Levinson (1977) Anesthesia for obstetrics. In: Miller RD, ed. Anesthesia 32, 517-523.
Siegel, J. (1999) The evolution of REM sleep. In Handbook of Behavioral State Control., Lydic R and Baghdoyan
eds., pp 87-100, CRC Press, Boca Raton.
Silverberg, D. and A. Oksenberg (1997) Essential hypertension and abnormal upper airway resistance during
sleep. Sleep 20, 794-806.
Teran-Santos, J., A. Jimenez-Gomez, J. Cordero-Guevara (1999) The association between sleep apnea and the
risk of traffic accidents. N Engl J Med 340, 847-851.
Thomason, J. and A. Russell (1986) Mechanical factors in the evolution of the mammalian secondary palate: a
theoretical analysis. Morphol 189(2),199-213.
Togias, A., R. Naclerio, and S. Peters et al. (1986) Local generation of sulfidopeptide leukotrienes upon nasal
provocation with cold, dry air. Am Rev Respir Dis 133, 1133-1137.
Utley, D., E. Shin, A. Clerck et al. (1997) A cost-effective and rational surgical approach to patients with snoring,
upper airway resistance syndrome, or obstructive sleep apnea syndrome. Laryngoscope 107, 726-734.
Veasey, S., P. Fenik, K. Panckeri, A. Pack and J. Hendricks (1999) The Effects of Trazodone with L-Tryptophan
on Sleep-disordered Breathing in the English Bulldog. Am J Respir Crit Care Med 160(5), 1659-1667.
Wark, K. and D. Richards (1999) Thermodynamics, 6th Ed., McGraw-Hill.
Wiegand, L., C. Zwillich, D. Wiegand and D. White (1991) Changes in upper airway muscle activation and
ventilation during phasic REM sleep in normal men. J Appl Physiol 71, 488-497.
Williams, G. and R. Nesse (1991) The Dawn of Darwinian Medicine. The Quarterly Review of Biology 66, 1-22.
Young, T., L. Evans, L. Finn and M. Palta (1997) Estimation of the clinically diagnosed proportion of sleep apnea
syndrome in middle-aged men and women. Sleep 20, 705-706.
Young, T., M. Palta, J. Dempsey, J. Skatrud, S. Weber, S. Badr (1993) The occurrence of sleep-disordered
breathing among middle-aged adults. N Engl J Med 32, 1230-1235.
Young, T., P. Peppard, M. Palta, K. Hla, L. Finn, B. Morgan and J. Skatrud (1997) Population-based study of
sleep-disordered breathing as a risk factor for hypertension. Arch Intern Med 157, 1746-1752.
Zielinski, J., A. Zgierska, M. Polakowska et al. (1999) Snoring and excessive daytime somnolence among Polish
middle-aged adults. Eur Respir J 14, 946-950.